Система свертываемости крови
Конспект из книги «Основы клинической гирудотерапии» Н.И. Сулим
Под термином «гемостаз» понимают комплекс реакций, направленный на остановку кровотечения при травме сосудов. В действительности, значение систем гемостаза намного сложнее и далеко выходит за рамки борьбы с кровотечениями. Основными задачами системы гемостаза являются сохранение жидкого состояния циркулирующей и депонированной крови, регуляция транскапиллярного обмена, резистентности сосудистой стенки, влияние на интенсивность репаративных процессов.
Принято различать: сосудисто-тромбоцитарный гемостаз и процесс свертывания крови. В первом случае речь идет об остановке кровотечения из мелких кровеносных сосудов с низким кровяным давлением, диаметр которых не превышает 100 мкм, во втором — о борьбе с кровопотерей при повреждении артерий и вен. Такое деление носит условный характер, ибо как при повреждении мелких, так и крупных кровеносных сосудов всегда наряду с образованием тромбоцитарной пробки осуществляется свертывание крови.
Вместе с тем, подобное разделение чрезвычайно удобно для клиницистов, ибо при нарушениях сосудисто-тромбоцитарного гемостаза прокол кожи пальца или мочки уха сопровождается длительным кровотечением, тогда как время свертывания крови остается в норме. При патологии свертывающей системы крови время кровотечения значительно не изменяется, хотя образование фибринового сгустка может не наступать часами, что, в частности, наблюдается при гемофилиях А и В.
Сосудисто-тромбоцитарный гемостаз
Сосудисто-тромбоцитарный гемостаз сводится к образованию тромбоцитарной пробки, или тромбоцитарного тромба.
Три стадии сосудисто-тромбоцитарного гемостаза
- временный (первичный и вторичный) спазм сосудов;
- образование тромбоцитарной пробки за счет адгезии (прикрепления к поврежденной поверхности) и агрегации (склеивания между собой) кровяных пластинок;
- ретракция (сокращение и уплотнение) тромбоцитарной пробки.
Временный спазм сосудов
Буквально через доли секунды после травмы наблюдается первичный спазм кровеносных сосудов, благодаря чему кровотечение в первый момент может не возникнуть или носит ограниченный характер. Первичный спазм сосудов обусловлен выбросом в кровь в ответ на болевое раздражение адреналина и норадреналина и длится не более 10-15 сек. В дальнейшем наступает вторичный спазм, обусловленный активацией тромбоцитов и отдачей в кровь сосудосуживающих агентов — серотонина, ТхА2, адреналина и др.
Первичная (обратимая) агрегация тромбоцитов
Повреждение сосудов сопровождается немедленной активацией тромбоцитов, что связано с появлением высоких концентраций АДФ (из разрушающихся эритроцитов и травмированных сосудов), а также обнажением субэндотелия, коллагеновых и фибриллярных структур. Начинается адгезия тромбоцитов к коллагену и другим адгезивным белкам субэндотелия.
При повреждении крупных артерий и вен, тромбоциты адгезируют непосредственно к обнаженным волокнам коллагена через коллагеновые рецепторы — GP-Ib-IIa.
При травме мелких артерий и артериол, прилипание тромбоцитов обусловлено наличием в плазме и кровяных пластинках, а также высвобождением из эндотелия особого белка — фактора фон Виллебранда (vWF), имеющего 3 активных центра, два из которых связываются с рецепторами тромбоцитов (GPIb), а один — с субэндотелием или коллагеновыми волокнами. Таким образом, тромбоцит с помощью vWF оказывается «подвешенным» к травмированной поверхности сосуда.
Из адгезирующих тромбоцитов, как и из поврежденного эндотелия, высвобождается АДФ, являющаяся важнейшим индуктором агрегации. Под влиянием АДФ тромбоциты прилипают к присоединившимся к эндотелию кровяным пластинкам, а также склеиваются между собой, образуя агрегаты, являющиеся основой тромбоцитарной пробки. Усилению агрегации способствуют фактор активации тромбоцитов (PAF), а также тромбин, всегда появляющийся в результате свертывания крови в зоне травмы.
Под воздействием слабых агонистов (АДФ, PAF, адреналин, серотонин, витронектин, фибронектин и др.) на мембране тромбоцитов начинается экспрессия рецепторов к фибриногену (GPIIb-IIIa). Благодаря им в присутствии ионов Са2+ фибриноген связывает между собой 2 близлежащие кровяные пластинки.
На этом этапе агрегация носит обратимый характер, ибо вслед за агрегацией может наступить частичный или полный распад агрегатов — дезагрегация. Более того, так как связь между тромбоцитами непрочна, то часть агрегатов может отрываться и уноситься током крови. Такая агрегация носит наименование первичной, или обратимой. Разумеется, первичная агрегация не способна остановить кровотечение даже из очень мелких кровеносных сосудов (капилляров, венул, артериол).
Ретракция сгустка
Более сложен механизм вторичной агрегации, сопровождающийся тромбоцитарной секрецией. Для завершения гемостаза требуется присоединение ряда дополнительных механизмов активации с включением обратных связей (обратной афферентации в пределах тромбоцита). Слабые агонисты приводят к поступлению сигнала внутрь кровяных пластинок, в результате чего в них увеличивается содержание цитоплазматического Са2+ и наступает активация фосфолипазы А2. Последняя приводит к освобождению из мембраны тромбоцита арахидоновой кислоты, которая в результате цикла последовательных реакций превращается в чрезвычайно активные соединения PgG2, PgH2 и тромбоксан А2 (ТхА2), являющиеся одновременно сильным агонистом агрегации и вазоконстриктором.
Выделяясь из тромбоцитов, PgG2, PgH2 и особенно ТхА2 осуществляют так называемую первую положительную связь, заключающуюся в усилении экспрессии фибриногеновых рецепторов, а также усиливают сигнал, передаваемый внутрь тромбоцита. При этом ТхА2, вызывает выделение ионов Са2+ из плотной тубулярной системы в цитоплазму, что способствует развитию финальных ферментных реакций систем гемостаза в самом тромбоците. К таким реакциям, прежде всего, относится активация актомиозиновой системы, а также фосфорилирование белков. Этот путь, начавшийся с активизации фосфолипазы С, завершается активацией протеинкиназы С с образованием инозилтрифосфата, способного, как и ТхА2, повышать уровень Са2+.
Комплекс перечисленных реакций ведет, в конечном счете, к сокращению актомиозина (тромбостенина) тромбоцитов, что сопровождается повышением внутриклеточного давления, приводящего к секреторным реакциям (реакция высвобождения) и сокращению тромбоцитарной пробки. При этом кровяные пластинки подтягиваются друг к другу, тромбоцитарная пробка не только сокращается, но и уплотняется, т.е. наступает ее ретракция.
Из тромбоцитов, подвергшихся адгезии и агрегации, усиленно секретируются гранулы и содержащиеся в них биологически активные продукты — АДФ, PAF, адреналин, норадреналин, фактор Р4, ТхА2, фибриноген, vWF, тромбоспондин, фибронектин, витронектин и многие другие. Все это значительно укрепляет тромбоцитарный тромб (рис. 1).
Рис. 1. Состав гранул тромбоцитов и их высвобождение под влиянием стимуляторов агрегации.
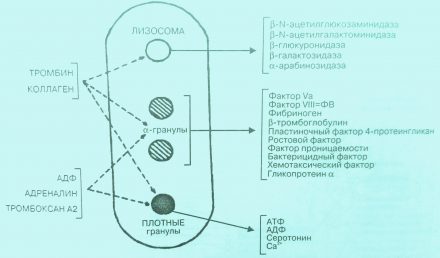
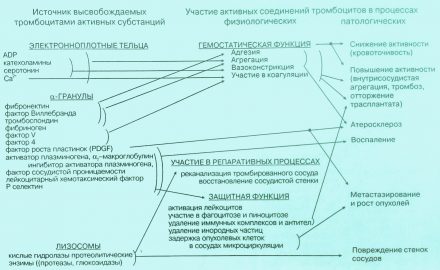
Следует обратить внимание на то, что из кровяных пластинок в процессе реакции высвобождения выделяется фактор роста, или иначе митогенный фактор, играющий важную роль в процессе репарации поврежденных стенок сосудов, а в условиях патологии способствующий развитию атеросклероза. Реканализации (восстановлению проходимости) сосуда способствуют лизосомальные энзимы, выделяемые из g-rpaнул (лизосом) (рис. 2).
Рис. 2. Продукты тромбоцитарной секреции в физиологических и патологических реакциях организма (по А.С. Шитиковой)
Одновременно с высвобождением тромбоцитарных факторов происходит образование тромбина, резко усиливающего агрегацию и приводящего к появлению сети фибрина в которой застревают отдельные эритроциты и лейкоциты.
Важно!!! В условиях нормы остановка кровотечения из мелких сосудов занимает от 2-х до 4-х минут.
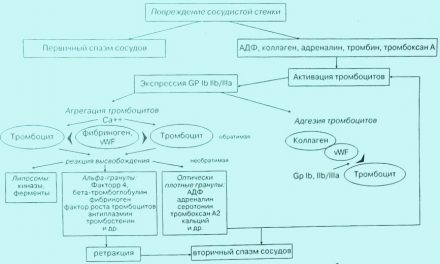
Общая схема сосудисто-тромбоцитарного гемостаза
Рис. 3. Схема сосудисто-тромбоцитарного гемостаза. Условные обозначения: АДФ — аденозиндифосфат, ГП — гликопротеины, КА — катехоламины vWF — фактор Виллибранда
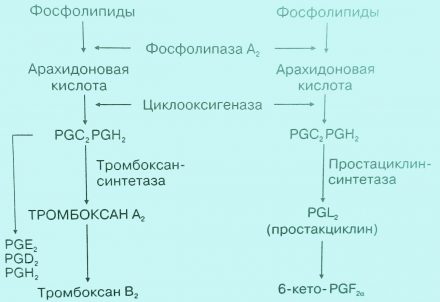
Роль простагландинов в сосудисто-тромбоцитарном гемостазе
Чрезвычайно важную роль в регуляции сосудисто-тромбоцитарного гемостаза играют производные арахидоновой кислоты — простагландин I2 (PgI2), или простациклин и ТхА2.
PgI2 образуется эндотелиальными клетками под влиянием фермента простациклинсинтетазы. В физиологических условиях действие PgI2 преобладает над ТхА2 — мощным агрегирующим агентом тромбоцитов. Вот почему в циркуляции у здорового человека агрегация тромбоцитов носит ограниченный характер.
При повреждении эндотелия в месте травмы образование PgI2 нарушается, в результате чего начинает преобладать действие ТхА2 и создаются благоприятные условия для агрегации тромбоцитов.
Аналогичная картина наблюдается при заболеваниях, сопровождающихся повреждением сосудистой стенки (эндотелиозы). В этих случаях в местах повреждения сосудов образуются так называемые белые тромбы, состоящие преимущественно из тромбоцитов. Наличие локальных повреждений коронарных сосудов является одной из ведущих причин возникновения стенокардии, инфаркта миокарда в результате обратимой (стенокардия) и необратимой (инфаркт) агрегации тромбоцитов с последующим цементированием тромбоцитарной пробки нитями фибрина.
Рис. 4. Схема, отражающая участие простагландинов в регуляции функции тромбоцитов
Процесс свертывания крови
При повреждении крупных кровеносных сосудов (артерий, вен) также происходит образование тромбоцитарной пробки, но она не способна остановить кровотечение, ибо легко вымывается током крови. Основное значение в этом процессе принадлежит свертыванию крови, сопровождающемуся в конечном итоге образованием плотного фибринового сгустка.
В настоящее время установлено, что свертывание крови является ферментативным процессом. Следует, однако, заметить, что основоположником ферментативной теории свертывания крови является отечественный ученый, профессор Дерптского университета А. А. Шмидт, опубликовавший с 1861 по 1895 год ряд работ, посвященных механизмам формирования фибринового сгустка. Эта теория лишь в начале XX века была поддержана немецким ученым Р. Моравитцем и получила общее признание.
Плазменные факторы свертывания крови
В свертывании крови принимает участие комплекс белков, находящихся в плазме (плазменные факторы гемокоагуляции), большинство из которых являются проферментами. В отличие от тромбоцитарных факторов, они обозначаются римскими цифрами (фактор I, II и т.д.).
Активация плазменных факторов происходит главным образом за счет протеолиза и сопровождается отщеплением пептидных ингибиторов. Для обозначения этого процесса к номеру фактора присоединяется буква «а» (фактор IIа, Va, VIIa и т.д.).
Плазменные факторы разделяются на две группы: витамин-К-зависимые, которые образуются преимущественно в печени при участии витамина К, и витамин-К-независимые, для синтеза которых витамин К не требуется. Такое разделение чрезвычайно удобно для клиники, ибо при угрозах внутрисосудистого тромбообразования врач может с помощью лекарственных препаратов нарушить синтез витамин-К-зависимых факторов и значительно снизить риск тромбоза (табл. 1).
Таблица 1. Плазменные факторы свертывания крови
|
Фактор |
Название фактора |
Свойства и функции |
| I | Фибриноген | Белок-гликопротеин. Образуется в печени. Под влиянием тромбина переходит в фибрин. Принимает участие в агрегации тромбоцитов. Необходим для репарации тканей. |
| II | Протромбин | Белок-гликопротеин. Неактивная форма фермента тромбина. Под влиянием протромбиназы переходит в тромбин (фактор IIa). Синтезируется в печени при участии витамина К. |
| III | Тромбопластин | Состоит из белка апопротеина III и комплекса фосфолипидов. Входит в состав мембран многих тканей. Является матрицей для развертывания реакций, направленных на образование протромбиназы по внешнему механизму. |
| IV | Кальций | Участвует в образовании комплексов, входящих в состав теназы и протромбиназы. Необходим для агрегации тромбоцитов, реакции высвобождения, ретракции. |
| V | Проакцелерин, Ас-глобулин |
Образуется в печени. Витамин-К-независим. Активируется тромбином. Входит в состав протромбиназного комплекса. |
| VI | Акцелерин | Потенцирует превращение протромбина в тромбин. |
| VII | Проконвертин | Синтезируется в печени при участии витамина К. Принимает участие в формировании протромбиназы по внешнему механизму. Активируется при взаимодействии с тромбопластином и факторами XIIa, Xa, IXa, IIa. |
| VIIIC | Антигемофильный глобулин А (АГГ) | Сложный гликопротеид. Место синтеза точно не установлено. В плазме образует комплекс с vWF и специфическим антигеном. Активируется тромбином. Входит в состав геназного комплекса. При его отсутствии или резком снижении возникает заболевание гемофилия А. |
| IX | Антигемофильный глобулин В, фактор Кристмаса |
Бета-глобулин, образуется в печени при участии витамина К. Активируется тромбином и фактором VIIa. Переводит фактор X в Xa. При его отсутствии или резком снижении возникает заболевание гемофилия В. |
| X | Тромботропин, фактор Стюарта-Прауэра |
Гликопротеид, вырабатывается в печени при участии витамина К. Фактор Xa является основной частью протромбиназного комплекса. Активируется факторами VIIа и IXа. Переводит фактор II в IIa. |
| XI | Предшественник плазменного тромбопластина, фактор Розенталя |
Гликопротеид. Активируется фактором XIIa, калликреином совместно с высокомоллекулярным кининогеном (ВМК). |
| XII | Фактор контактной активации, фактор Хагемана |
Белок. Активируется отрицательно заряженными поверхностями, адреналином, калликреином. Запускает внешний и внутренний механизм образования протромбиназы и фибринолиза, активирует фактор XI и прекалликреин. |
| XIII | Фибринстабилизирующий фактор (ФСФ), фибриназа |
Глобулин. Синтезируется фибробластами и мегакариоцитами. Стабилизирует фибрин. Необходим для нормального течения репаративных процессов. |
| Фактор Флетчер, плазменный прекалликреин |
Белок. Активирует факторы XII, плазминоген и ВМК. | |
| Фактор Фитцжеральда, высокомолекулярный кининоген (ВМК) |
Активируется калликреином, принимает участие в активации фактора XII, XI и фибринолизе. | |
| Фактор Виллебранда | Компонент фактора VIII, вырабатывается в эндотелии, в кровотоке, соединяясь с коагуляционной частью, образует полиоценный фактор VIII (антигемофильный глобулин А). |
Эритроцитарные факторы свертывания крови
В эритроцитах обнаружен ряд соединений, аналогичных тромбоцитарным факторам. Наиважнейшим из них является частичный тромбопластин, или фосфолипидный фактор (напоминает фактор Р3), который входит в состав мембраны. Кроме того, эритроциты содержат антигепариновый фактор, большое количество АДФ, фибриназу и другие соединения, имеющие отношение к гемостазу. При травме сосуда около 1% наименее стойких эритроцитов вытекающей крови разрушается, что способствует образованию тромбоцитарной пробки и фибринового сгустка.
Особенно велика роль эритроцитов в свертывании крови при их массовом разрушении, что наблюдается при переливании несовместимой крови, резус-конфликте матери и плода и гемолитических анемиях.
Лейкоцитарные факторы свертывания крови
Лейкоциты содержат факторы свертывания, получившие наименование лейкоцитарных. В частности, моноциты и макрофаги при стимуляции Аг синтезируют белковую часть тромбопластина — апопротеин III (тканевой фактор), что значительно ускоряет свертывание крови. Эти же клетки являются продуцентами витамин-К-зависимых факторов свертывания — IX, VII и X. Приведенные факты являются одной из основных причин возникновения диссеминированного (распространенного) внутрисосудистого свертывания крови (или ДВС-синдрома) при многих воспалительных и инфекционных заболеваниях, что значительно отягощает течение патологического процесса, а иногда служит причиной смерти больных.
Тканевые факторы свертывания крови
Важная роль в процессе свертывания крови отводится тканевым факторам, к которым в первую очередь относится тромбопластин (фактор III, тканевой фактор — TF). TF состоит из белковой части — апопротеина III и комплекса фосфолипидов — и нередко представляет собой отломок клеточных мембран. Большая часть TF экспонирована наружу и включает 2 структурных домена. При разрушении тканей или стимуляции эндотелия эндотоксином и провоспалительными цитокинами TF способен поступать в кровоток и вызывать развитие ДВС-синдрома.
Механизм свертывания крови
Процесс свертывания крови представляет собой ферментный каскад, в котором проферменты, переходя в активное состояние (сериновые протеиназы), способны активировать другие факторы свертывания крови. Подобная активация может носить последовательный и ретроградный характер. При этом активация факторов свертывания осуществляется за счет протеолиза, приводящего к перестройке молекул и отщеплению пептидов, обладающих слабым антикоагулянтным действием.
Процесс свертывания крови может быть разделен на 3 фазы
- комплекс последовательных реакций, приводящих к образованию протромбиназы;
- переход протромбина в тромбин (фактора II в фактор IIа);
- из фибриногена образуется фибриновый сгусток.
Образование протромбиназы
Образование протромбиназы может осуществляться по внешнему и внутреннему механизму. Внешний механизм предполагает обязательное присутствие тромбопластина (TF, или F-III), внутренний же связан с участием тромбоцитов (парциальный тромбопластин, или фактор Р3). Вместе с тем, внутренний и внешний пути образования протромбиназы имеют много общего, ибо активируются одними и теми же факторами (фактор ХIIа, калликреин, ВМК и др.), а также приводят в конечном итоге к появлению одного и того же активного фермента — фактора Ха, выполняющего в комплексе с фактором Va функции протромбиназы. При этом как полный, так и парциальный тромбопластин служат матрицами, на которых развертывается цикл ферментативных реакций.
Важная роль в процессе свертывания крови отводится глицерофосфолипидам и, в частности, фосфатидилсерину и фосфатидилэтаноламину в бислое мембраны. Одной из особенностей бислоя является его асимметрия. В наружном листке бислойной мембраны, контрактирующей с кровью, преобладают в основном фосфатидилхолин и сфингомиелин. Как известно, эти фосфолипиды содержат фосфохолин, обеспечивающий атромбогенность мембран. Молекула этих фосфолипидов электронейтральна — в ней нет преобладания одного из зарядов.
Фосфатидилсерин и фосфатидилэтаноламин расположены преимущественно во внутреннем слое мембраны. Головка указанных фосфолипидов несет два отрицательных заряда и один положительный, т.е. на ней преобладает отрицательный заряд. Инициация свертывания крови может наступить лишь тогда, когда эти фосфолипиды появятся на наружной поверхности мембраны.
Из сказанного вытекает, что для инициации свертывания крови необходимо нарушить исходную асимметрию фосфолипидов мембраны, что может произойти только за счет обмена фосфолипидов между слоями, или, иначе, флип-флопа. Как это происходит при повреждении кровеносного сосуда?
Мы уже отмечали, что по обе стороны мембраны существует ионная асимметрия. Для процесса свертывания крови очень важна асимметрия в содержании ионов Са2+, концентрация которого в плазме и интерстициальной жидкости в десять тысяч раз больше, чем в цитоплазме клетки и тромбоците. Как только травмируется стенка сосуда, в цитоплазму из внеклеточной жидкости или из внутриклеточного депо переходит значительное количество ионов Са2+. Поступление Са2+ в тромбоцит или клетки (травмированный эндотелий и т.п.) разрыхляет мембрану и включает механизмы поддержания асимметрии фосфолипидного бислоя. При этом молекулы фосфатидилсерина и фосфатидилэтаноламина, несущие суммарные отрицательные заряды, переходят на поверхность мембраны.
Почему же нарушается асимметрия в содержании отдельных фосфолипидов в наружном и внутреннем слоях мембраны? Недавно появился ряд сообщений о том, что зависимый от энергии процесс концентрации аминофосфолипидов преимущественно во внутреннем листке мембраны связан с функционированием специфичных синергично действующих трансмембранных белков-переносчиков — транслоказ.
Аминофосфолипидные транслоказы осуществляют однонаправленное передвижение фосфатидилсерина и фосфатиднлэтаноламина во внутренний листок мембраны. При активации клеток, в том числе кровяных пластинок, при повышении уровня цитоплазматического Са2+, при уменьшении концентрации АТФ и при ряде других сдвигов происходит ингибиция транслоказ. При этом наступает двунаправленное трансмембранное перемещение всех мембранных фосфолипидов, приводящее к значительному выравниванию их концентрации в обоих листках мембраны.
Но как только на поверхности клеточной мембраны увеличивается концентрация отрицательно заряженных фосфолипидов и они входят в соприкосновение с кровью, содержащей громадную концентрацию ионов Са2, то образуются кластеры — активные зоны, к которым прикрепляются факторы свертывания. При этом ионы Са2+ выполняют следующие функции:
1. Они необходимы для конформации факторов свертывания, после чего последние способны принимать участие в ферментативных реакциях гемостаза.
2. Они являются связующими мостиками между белковыми компонентами и клеточными мембранами. Эти реакции осуществляются следующим образом: ионы Са2+, с одной стороны, присоединяются к головкам фосфатидилсерина, а с другой — соединяются с остатками g-карбоксиглутаминовой кислоты, которая входит в состав ряда факторов свертывания крови (V, VIII, IX и др.). За счет таких кальциевых мостиков происходит первоначальное ориентирование на фосфолипидной поверхности факторов свертывания крови, и в результате конформации белковых молекул открываются активные центры.
Без ионов Са2+ не может происходить образование кластеров и не осуществляется взаимодействие друг с другом ферментов, участвующих в свертывании крови.
Формирование протромбиназы по внешнему пути начинается с активации фактора VII при его взаимодействия с тромбопластином‚ а также с факторами XIIа, IXа, Ха и калликреином. В свою очередь, фактор VIIa активирует не только фактор Х, но и IX. B процессе образования протромбиназы по внешнему механизму могут также принимать участие факторы IХа и VIIIa, образующие активный комплекс на фосфолипидной матрице. Однако эта реакция протекает относительно медленно.
Формирование протромбиназы по внешнему пути происходит чрезвычайно быстро (занимает секунды) и ведет к появлению фактора Ха и небольших порций тромбина (IIa), который способствует необратимой агрегации тромбоцитов, активации факторов VIII и V и значительно о ускоряет образование протромбиназы по внутреннему и внешнему механизмам.
Инициатором внутреннего пути образования протромбиназы является фактор XII, который активируется травмированной поверхностью, кожей, коллагеном, адреналином, после чего переводит фактор XI в XIа.
В этой реакции принимает участие калликреин (активируется фактором ХIIа) и ВМК (активируется калликреином).
Фактор ХIа оказывает непосредственное влияние на фактор IX, переводя его в фактор IXa. Специфическая деятельность последнего направлена на протеолиз фактора X (перевод его в фактор Ха) и протекает на поверхности фосфолипидов тромбоцита при обязательном участии фактора VIII (или VIIIa). Комплекс факторов IXa, VIIIa на фосфолипидной поверхности тромбоцитов получил наименование теназы, или теназного комплекса.
Как уже отмечалось, в процессе свертывания крови принимают участие прекалликреин и ВМК, благодаря которым (как и фактору XII) происходит объединение внешнего и внутреннего путей свертывания крови. В настоящее время установлено, что при травме сосуда всегда происходит освобождение металлопротеидов, переводящих прекалликреин в калликреин. Под воздействием калликреина ВМК переходит в ВМКа. Кроме того, калликреин способствует активации факторов VII и XII, что также сопровождается запуском каскадного механизма свертывания крови.
Переход протромбина в тромбин
Вторая фаза процесса свертывания крови (переход фактора II в фактор IIа) осуществляется под влиянием протромбиназы (комплекса Xa+Va+Са2+) и сводится к протеолитическому расщеплению протромбина, благодаря чему появляется фермент тромбин, обладающий свертывающей активностью.
Переход фибриногена в фибрин
Третья стадия процесса свертывания крови — переход фибриногена в фибрин — включает 3 этапа. На первом из них под влиянием фактора IIа от фибриногена отщепляются 2 фибринпептида А и 2 фибринпептида В, в результате чего образуются фибрин-мономеры. На втором этапе, благодаря процессу полимеризации, формируются вначале димеры и олигомеры фибрина, трансформирующиеся в дальнейшем в волокна фибрина — протофибриллы легкорастворимого фибрина, или фибрина s (soluble), быстро лизирующегося под влиянием протеаз (плазмина, трипсина). В процесс образования фибрина вмешивается фактор XIII (фибриназа, фибринстабилизирующий фактор), который после активации тромбином в присутствии Са2+ прошивает фибринполимеры дополнительными перекрестными связями, благодаря чему появляется труднорастворимый фибрин, или фибрин i (insoluble). В результате этой реакции сгусток становится резистентным к мочевине и фибринолитическим (протеолитическим) агентам и плохо поддается разрушению.
Рис. 5. Схема свертывания крови. Условные обозначения: тонкие стрелки — активация, толстые стрелки — переход фактора в активное состояние, ВМК — высокомолекулярный кининоген, I — фибриноген, Im — фибринмономер, Is — легкорастворимый фибрин, Ii — труднорастворимый фибрин.
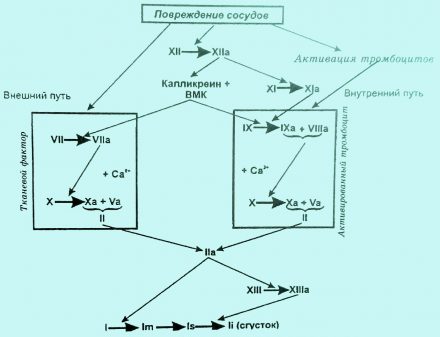
Образовавшийся фибриновый сгусток, благодаря тромбоцитам, входящим в его структуру, сокращается и уплотняется (наступает ретракция) и прочно закупоривает поврежденный сосуд.
Естественные антикоагулянты
Несмотря на то что в циркуляции имеются все факторы, необходимые для образования тромба, в естественных условиях при наличии целых сосудов кровь остается жидкой. Это обусловлено наличием в кровотоке противосвертывающих веществ, получивших название естественные антикоагулянты, и фибринолитического звена системы гемостаза.
Естественные антикоагулянты делятся на первичные и вторичные. Первичные антикоагулянты всегда присутствуют в циркуляции, вторичные — образуются в результате протеолитического расщепления факторов свертывания крови в процессе формирования и растворения фибринового сгустка.
Первичные антикоагулянты можно разделить на 3 основные группы: 1) обладающие антитромбопластическим и антипротромбиназным действием (антитромбопластины); 2) связывающие тромбин (антитромбины); 3) предупреждающие переход фибриногена в фибрин (ингибиторы самосборки фибрина).
К антитромбопластинам, в первую очередь, относится ингибитор внешнего пути свертывания (TFPI). Установлено, что он способен блокировать комплекс факторов III+VII+Ха, благодаря чему предотвращается образование протромбиназы по внешнему манизму. Недавно обнаружен еще один ингибятФ внешнего пути образования протромбиназы, получивший наименование TFPI-2 (анексин V), однако он обладает меньшей активностью, чем TFPI.
К ингибиторам, блокирующим образование протромбиназы, относятся витамин-К-зависимые протеины С, S (РrС, PrS) и особый белок, синтезируемый эндотелием, — тромбомодулин. Под воздействием тромбомодулина и связанного с ним тромбина РrС переходит в активное состояние (Рrа), чему способствует кофактор PrS, РrСа разрезает пополам факторы V и VIII и тем самым препятствует образованию протромбиназы внутреннему пути и переходу протромбина в тромбин.
Недавно появились сообщения, что PrS способен связывать фактор Ха. Эта реакция не зависит от фосфолипидной поверхности и усиливается в присутствии РrС.
Одним из ведущих антикоагулянтов является белок антитромбин III (A-III), имеющий молекулярную массу (ММ) 58 кД. Самостоятельно А-III обладает слабым антикоагулянтным действием. В то же время он способен образовывать комплекс с сульфатированным полисахаридом гликозамингликаном гепарином (Г) — А-III+Г. Этот комплекс связывает факторы IIа, IXa, Ха, ХIа, ХIIа, калликреин и плазмин. Существует высокомолекулярный гепарин (нефракдионированный) с ММ от 25 до 35 кД и низкомолекулярный гепарин с ММ менее 5 кД. Последний в меньшей степени нуждается во взаимодействии с А-III и нейтрализует преимущественно фактор Ха, ибо его цепочка мала и «не дотягивается» до тромбина. Низкомолекулярный Г в большей степени, чем высокомолекулярный, способствует высвобождению из эндотелия TFPI, благодаря чему его антикоагулянтная активность возрастает. Следует также заметить, что низкомолекулярные гепарины ингибируют прокоагулянтную активность поврежденного эндотелия и некоторых протеаз, выделяемых гранулоцитами и макрофагами (рис. 6).
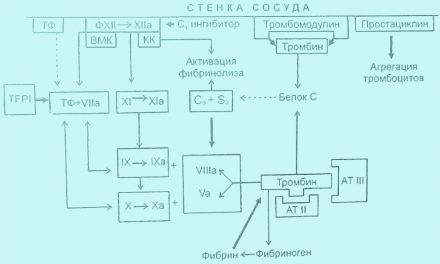
За последнее время появились сообщения о наличии еще одного антикоагулянта — белка антитромбина II, однако его активность уступает А-III. Важным ингибитором свертывания является кофактор гепарина II, связывающего тромбин. Его действие усиливается во много раз при взаимодействии с гепарином.
Ингибитором тромбина, факторов IXa, XIa, ХIIа и плазмина является a1-антитрипсин. Слабым ингибитором тромбина, калликреина и плазмина служит а2-макроглобулин.
К первичным антикоагулянтам следует также отнести аутоантитела к активным факторам свертывания крови (IIа, Ха и др.), которые всегда присутствуют в кровотоке, а также покинувшие клетку рецепторы (так называемые «плавающие» рецепторы) к активированным факторам свертывания крови. Однако их роль в условиях нормы и патологии пока еще далека от окончательного выяснения.
Следует заметить, что при снижении концентрации первичных естественных антикоагулянтов создаются благоприятные условия для развития тромбофилий и диссеминированного внутрисосудистого свертывания крови — ДВС-синдрома.
Таблица 2. Основные естественные антикоагулянты (первичные)
| Антитромбин III | Альфа2-глобулин. Синтезируется в печени. Прогрессивно действующий ингибитор тромбина, факторов IXа, Xа, XIа, XIIа, калликреина и в меньшей степени — плазмина и трипсина. Плазменный кофактор гепарина. |
| Гепарин | Сульфатированный полисахарид. Трансформирует антитромбин III из прогрессивного в антикоагулянт немедленного действия, значительно повышая его активность. Образует комплексы с тромбогенными белками и гормонами, обладающие антикоагулянтным и фибринолитическим действием. |
| Кофактор гепарина II | Слабый антикоагулянт, действующий в присутствии гепарина. |
| Альфа2-антиплазмин | Белок. Ингибирует действие плазмина, трипсина, хемотрипсина, калликреина, фактора Xа, урокиназы. |
| Альфа2-макроглобулин | Слабый прогрессивный ингибитор тромбина, калликреина, плазмина и трипсина. |
| Альфа1-антитрипсин | Ингибитор тромбина, факторов IXа, XIа, XIIа, трипсина и плазмина. |
| С1-эстеразный ингибитор, или ингибитор комплимента I | Альфа1-нейроаминогликопротеид. Инактивирует калликреин, предотвращая его действие на кининоген, факторы XIIа, IXа, XIа и плазмин. |
| TFPI | Ингибирует комплекс TF+VII+Xа. |
| TFPI-2 или анексин V | Образуется в плаценте. Ингибирует комплекс TF+VII+Xа. |
| Протеин С | Витамин-К-зависимый белок. Образуется в печени и эндотелии. Обладает свойствами сериновой протеазы. Инактивирует факторы Va и VIIIа и стимулирует фибринолиз. |
| Протеин S | Витамин-К-зависимый белок. Образуется эндотелиальными клетками. Усиливает действие протеина С. |
| Тромбомодулин | Гликопротеин, фиксированный на цитоплазматической мембране эндотелия. Кофактор протеина С, связывается с фактором IIa и инактивирует его. |
| Ингибитор самосборки фибрина | Полипептид, образуется в различных тканях. Действует на фибрин мономер и полимер. |
| Плавающие рецепторы | Гликопротеиды, связывающие факторы IIa и Ха, а возможно, и другие сериновые протеазы |
| Аутоантитела к активным факторам свертывания | Находятся в плазме, ингибируют факторы и др. |
К вторичным антикоагулянтам относятся «отработанные» факторы свертывания крови (принявшие участие в свертывании) и продукты деградации фибриногена и фибрина (ПДФ), обладающие антиагрегационным и противосвертывающим действием, а также стимулирующие фибринолиз. Роль вторичных антикоагулянтов сводится к ограничению внутрисосудистого свертывания крови и распространения тромба по сосудам.
Фибринолиз
Фибринолиз является неотъемлемой частью системы гемостаза, всегда сопровождает процесс свертывания крови и даже активируется теми же самыми факторами (ХIIа, калликреином, ВМК и др.). Являясь важной защитной реакцией, фибринолиз предотвращает закупорку кровеносных сосудов фибриновыми сгустками, а также приводит к реканализации сосудов после остановки кровотечения. Компоненты фибринолиза играют важную роль в удалении внеклеточного матрикса и, кроме того, регулируют рост и деление клеток, заживление ран, регенерацию мышц, рост и метастазирование опухолей и т.д.
Ферментом, разрушающим фибрин, является плазмин (иногда его называют фибринолизин), который в циркуляции находится в неактивном состоянии в виде профермента плазминогена. Под воздействием его активаторов происходит расщепление пептидной связи Arg561-Val562 плазминогена, в результате чего образуется плазмин. Активный центр плазмина находится в легкой цепи, представляющей малоспецифичную протеазу, способную расщеплять практически все белки плазмы.
В кровотоке плазминоген встречается в двух основных формах: в виде нативного профермента с NH2-терминальной глутаминовой кислотой — глу-плазминогена, и в виде частично подвергшегося протеолизу — лиз-плазминогена. Последний приблизительно в 20 раз быстрее трансформируется физиологическими активаторами в плазмин, а также имеет большее сродство к фибрину.
Фибринолиз, как и процесс свертывания крови, может протекать по внешнему и внутреннему путям.
Внешний путь активации плазминогена
Внешний путь активации плазминогена осуществляется при участии тканевых активаторов, которые синтезируются главным образом в эндотелии. К ним, в первую очередь, относится тканевой активатор плазминогена (TPА).
Кроме того, активатором плазминогена является урокиназа, образуемая в почках (в юкстагломерулярном аппарате), а также фибробластами, эпителиальными клетками, пневмоцитами, децедуальными клетками плаценты и эндотелиоцитами. Многие клетки содержат рецепторы к урокиназе, что послужило основанием считать ее основным активатором фибринолиза в межклеточном пространстве, обеспечивающем протеолиз в процессе клеточного роста, деления и миграции клеток.
По мнению З.С. Баркагана, во внешнем пути активации фибринолиза принимают также участие активаторы форменных элементов крови — лейкоцитов, тромбоцитов и эритроцитов.
Внутренний путь активации фибринолиза
Внутренний путь активации фибринолиза, осуществляемый плазменными активаторами, разделяется на Хагеманзависимый и Хагеманнезависимый.
Хагеманзависимый фибринолиз осуществляется наиболее быстро и носит срочный характер. Его основное назначение сводится к очищению сосудистого русла от фибриновых сгустков, образующихся в процессе внутрисосудистого свертывания крови. Хагеманзависимый фибринолиз протекает под влиянием факторов ХIIа, калликреина и ВМК, которые переводят плазминоген в плазмин.
Хагеманнезависимый фибринолиз может осуществляться под влиянием протеинов С и S (рис. 7).
Рис. 7. Схема фибринолиза.
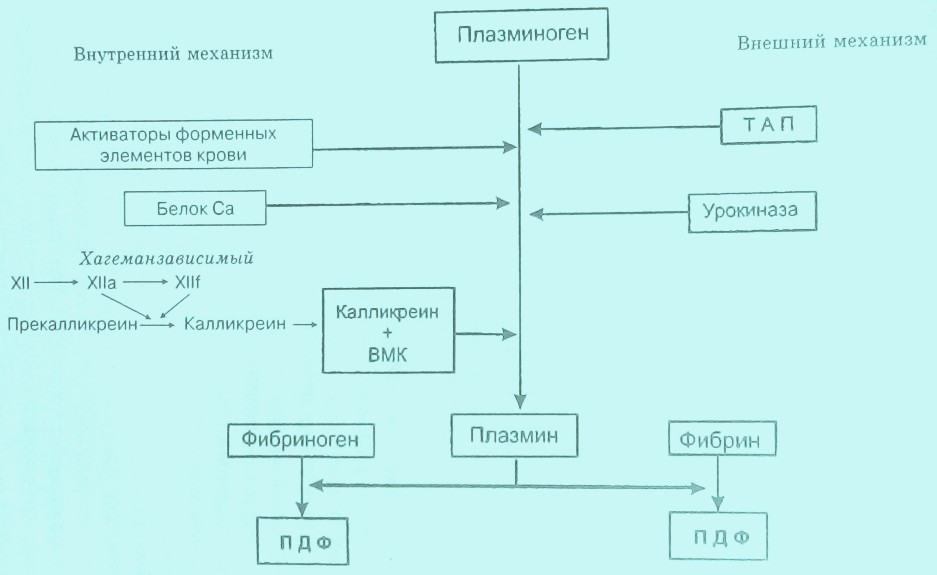
Образовавшийся в результате активации плазмин вызывает расщепление фибрина. При этом появляются ранние (крупномолекулярные) и поздние (низкомолекулярные) продукты деградации фибрина, или ПДФ.
Ингибиторы фибринолиза
До 90% всей антифибринолитической активности сосредоточено в а-гранулах тромбоцитов, которые выбрасываются в кровоток при их активации. В плазме находятся и ингибиторы фибринолиза. В настоящее время выявлено 4 типа ингибитора активатора плазминогена и урокиназы.
Важнейшим из них является ингибитор первого типа (PAI-1), который нередко называют эндотелиальным. Вместе с тем, он синтезируется не только эндотелием, но и гепатоцитами, моноцитами, макрофагами, фибробластами и мышечными клетками. Скапливаясь в местах повреждения эндотелия, тромбоциты также высвобождают PAI-1. PAI-1 является ингибитором сериновых протеаз. Его особенность заключается в том, что переход из неактивной в активную форму осуществляется без частичного протеолиза (за счет кон-формации молекулы) и является обратимым процессом. Хотя концентрация PAI-1 примерно в 1000 раз ниже, чем других ингибиторов протеаз, ему принадлежит основная роль в регуляции начальных стадий фибринолиза.
Важнейшим ингибитором фибринолиза является а2-антиплазмин, связывающий не только плазмин, но и трипсин, калликреин, урокиназу, ТАР и, следовательно, вмешивающийся как на ранних, так и на поздних стадиях фибринолиза.
Сильным ингибитором плазмина служит a1-протеазный ингибитор (a1-антитрипсин).
Кроме того, фибринолиз тормозится а2-макроглобулином, C1-эстеразным ингибитором, а также целым рядом ингибиторов активатора плазминогена, синтезируемых эндотелием, макрофагами, моноцитами и фибробластами.
Фибринолитическая активность крови во многом определяется соотношением активаторов и ингибиторов фибринолиза.
При ускорении свертывания крови и одновременном торможении фибринолиза создаются благоприятные условия для развития тромбозов, эмболии и ДВС-синдрома.
Наряду с ферментативным фибринолизом, по мнению профессора Б.А. Кудряшова и его учеников, существует так называемый неферментативный фибринолиз, который обусловлен комплексными соединениями естественного антикоагулянта гепарина с ферментами и гормонами. Неферментативный фибринолиз приводит к расщеплению нестабилизированного фибрина, очищая сосудистое русло от фибринмономеров и фибрина s.
Четыре уровня регуляции сосудисто-тромбоцитарного гемостаза, свертывания крови и фибринолиза
Свертывание крови, контактирующей со стеклом, травмированной поверхностью или кожей, осуществляется за 5-10 минут. Основное время в этом процессе уходит на образование протромбиназы, тогда как переход протромбина в тромбин и фибриногена в фибрин осуществляется довольно быстро. В естественных условиях время свертывания крови может уменьшаться (развивается гиперкоагуляция) или удлиняться (возникает гипокоагуляция).
Между тем образование тромбоцитарной пробки и остановка кровотечения из мелких сосудов осуществляется в течение 2-4 минут.
Млекулярный уровень регуляции
Молекулярный — предполагает поддержание гомеостатического баланса отдельных факторов, влияющих на сосудисто-тромбоцитарный гемостаз, свертывание крови и фибринолиз. При этом избыток фактора, возникающий по той или иной причине в организме, должен быть в кратчайшие сроки ликвидирован. Такой баланс постоянно поддерживается между простациклином (Pgl2) и ТхА2, прокоагулянтами и антикоагулянтами, активаторами и ингибиторами плазминогена.
Наличие клеточных рецепторов ко многим факторам свертывания крови и фибринолиза лежит в основе гомеостатического баланса в системе гемостаза на молекулярном уровне. Отрывающиеся от клетки рецепторы к факторам свертывания и фибринолиза («плавающие» рецепторы) приобретают новые свойства, становясь естественными антикоагулянтами, ингибиторами плазмина и активатора плазминогена.
Молекулярный уровень регуляции может осуществлять иммунная система с помощью образования Ат к активированными факторам свертывания крови и фибринолиза — IIа, Ха, ТАП и другим.
Необходимо также помнить, что существует генетический контроль над продукцией факторов, обеспечивающих образование и растворение кровяного сгустка.
Клеточный уровень регуляции
В кровотоке происходит постоянное потребление факторов свертывания и фибринолиза, что неминуемо должно приводить к восстановлению их концентрации. Этот процесс должен быть обусловлен или активированными факторами, или (что более вероятно) продуктами их распада. Если это так, то клетки, продуцирующие факторы свертывания и фибринолиза, должны нести на себе рецепторы к указанным соединениям или их депозитам. Такие рецепторы обнаружены на многих клетках к тромбину, калликреину, активатору плазминогена, плазмину, стрептокиназе, ПДФ и многим другим. Клеточная регуляция должна осуществляться по механизму обратной связи (обратной афферентации). Клеточный уровень регуляции систем гемостаза частично обеспечивается за счет «пристеночного» фибринолиза, возникающего при отложении фибрина на эндотелии сосудистой стенки.
Органный уровень регуляции
Органный уровень регуляции — обеспечивает оптимальные условия функционирования системы гемостаза в различных участках сосудистого русла. Благодаря этому уровню проявляется мозаичность сосудисто-тромбоцитарного гемостаза, свертывания крови и фибринолиза.
Нервно-гуморальная регуляция
Нервно-гуморальная регуляция контролирует состояние системы гемостаза от молекулярного до органного уровня, обеспечивая целостность реакции на уровне организма, главным образом, через симпатический и парасимпатический отделы вегетативной нервной системы, а также гормоны и различные биологически активные соединения.
Установлено, что при острой кровопотере, гипоксии, интенсивной мышечной работе, болевом раздражении, стрессе свертывание крови значительно ускоряется, что может привести к появлению фибрин-мономеров и даже фибрина s в сосудистом русле. Однако, благодаря одновременной активации фибринолиза, носящего защитный характер, появляющиеся сгустки фибрина быстро растворяются и не наносят вреда здоровому организму.
Ускорение свертывания крови и усиление фибринолиза при всех перечисленных состояниях связано с повышением тонуса симпатического отдела вегетативной нервной системы и поступлением в кровоток адреналина и норадреналина. При этом активируется фактор Хагемана, что приводит к запуску внешнего и внутреннего механизма образования протромбиназы, а также стимуляции Хагеман-зависимого фибринолиза. Кроме того, под влиянием адреналина усиливается образование апопротеина III — составной части тромбопластина — и наблюдается отрыв от эндотелия клеточных мембран, обладающих свойствами тромбопластина, что способствует резкому ускорению свертывания крови. Из эндотелия также выделяются ТАР и урокиназа, приводящие к стимуляции фибринолиза.
При повышении тонуса парасимпатического отдела вегетативной нервной систем (раздражение блуждающего нерва, введение ацетилхолина, пилокарпина) также наблюдается ускорение свертывания крови и стимуляция фибринолиза. Как это ни покажется на первый взгляд странным, но и в этих условиях происходит выброс тромбопластина и активаторов плазминогена из эндотелия сердца и сосудов.
Оказалось, что как сосудосуживающие, так и сосудорасширяющие воздействия вызывают со стороны свертывания крови и фибринолиза однотипный эффект — освобождение тканевого фактора и ТАР. Следовательно, основным эфферентным регулятором свертывания крови и фибринолиза является сосудистая стенка. Напомним также, что в эндотелии сосудов синтезируется Pgl2, препятствующий в кровотоке адгезии и агрегации тромбоцитов.
Вместе с тем, развивающаяся гиперкоагуляция может смениться гипокоагуляцией, которая носит в естественных условиях вторичный характер и обусловлена расходом (потреблением) тромбоцитов и плазменных факторов свертывания крови, образованием вторичных антикоагулянтов, а также рефлекторным выбросом в сосудистое русло гепарина и А-III в ответ на появление тромбина.
Важно!!! Следует отметить, что существует корковая регуляция системы гемостаза, что было блестяще доказано школами профессора Е.С. Иваницкого-Василенко и академика А.А. Маркосяна. В этих лабораториях были выработаны условные рефлексы как на ускорение, так и на замедление свертывания крови.